


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:我爱生化 作者:Robert Young

在生化世界里,我们常把 DNA 序列视为生命的“硬编码”,但越来越多的证据表明,基因组之外还有一层“软记忆”——表观遗传。“我爱生化”已发表多篇与“表观遗传”有关的推送(突破孟德尔定律!小鼠DNA甲基化呈现广泛非孟德尔遗传,新研究刷新我们对表观遗传继承的认知)(千万不要抽烟!新研究显示:青少年时期吸烟的男人会将有害的表观遗传特征传给未来的孩子,增加他们患哮喘、肥胖和肺功能低下的风险!)(新研究揭示了植物如何将对高浓度二氧化碳的反应“记忆”传递给它们的后代的表观遗传机制!)(科学家第一次实现了甲基化编辑的小鼠的跨代表观遗传,但究竟如何遗传下来的分子机制还是一个不解之谜!)!这种遗传方式不改写碱基序列,却能记录环境经历,并跨代传递。它正悄然重塑我们对生命进化的理解:达尔文的进化论即“基因随机突变”加上“自然选择”,真的就能解释一切吗?被主流科学打入冷宫 的200 多年前的法国博物学家Jean-Baptiste Lamarck(让-巴蒂斯特·拉马克)提出的“Inheritance of acquired traits(获得性状遗传)”真的就是一无是处吗?
今天写的这篇推送,我想从历史争议讲起,系统梳理线虫、果蝇、小鼠、植物、人类到原核生物的代表性案例,深入分子机制,并探讨对生化研究、医学和农业的启示。请准备好一大杯咖啡☕️,一起探索生命的“可遗传记忆”吧!
一、拉马克 vs 达尔文:一场关于“获得性”的世纪之争
拉马克在 1809 年Philosophie Zoologique(动物学哲学)中提出两大核心观点:一是用进废退。频繁使用的器官会发达,不用的会退化;二是获得性状遗传。个体在环境中获得的适应性变化,如长颈鹿🦒努力伸脖子吃高树叶,脖子变长,可以遗传给后代。

达尔文在1859 年On the Origin of Species(物种起源)中则强调随机变异和自然选择。20 世纪初,魏斯曼提出“种质-体质”分离(Weismann barrier):体细胞(soma)的变化不会影响生殖细胞(germline),获得性状因此无法遗传。这成为现代综合进化论的基石,拉马克学说被视为“经典错误”。
但科学从不停止。表观遗传学(Epigenetics)的兴起,正在重塑我们对“遗传”的理解。
二、表观遗传:DNA 序列之外的“可遗传记忆”
表观遗传强调不改变 DNA 碱基序列,但能影响基因表达的可遗传变化。主要机制包括:
(1)DNA 甲基化。最经典,主要发生在 CpG 二核苷酸序列岛(CpG islands)中的C上。甲基化通常使染色质致密化,阻碍转录因子结合或招募抑制蛋白,导致基因沉默。但若发生在基因体内,可以激活基因的表达。
甲基化可发生在印记基因上,实现父源或母源等位基因的单等位基因表达。
环境因素(如营养、压力、毒物)可改变甲基化模式,这些变化部分可通过生殖细胞传递给后代(跨代表观遗传)。
DNA 甲基化是表观遗传中最稳定的标记之一,可在细胞分裂中通过 DNMT1(维护甲基转移酶)忠实复制。
(2)组蛋白修饰。组蛋白尾部(尤其是 H3、H4)的氨基酸残基(特别是K和R)可发生多种化学修饰,如乙酰化、甲基化、泛素化、乳酰化、多巴胺化等。
乙酰化通常激活基因表达,使染色质松散开放;甲基化具有双重作用,如H3K4me3激活,H3K27me3则抑制。
组蛋白修饰构成“组蛋白密码”(Histone Code),不同组合决定基因的“开”或“关”。这些修饰可在细胞分裂中部分保留,实现细胞记忆;在生殖细胞中也可跨代传递(如应激诱导的组蛋白标记变化)。
(3)非编码 RNA的作用。如miRNA、siRNA、piRNA、lncRNA、circRNA 等基因表达的调控。
非编码 RNA 是“移动信号”,可将环境信息从 体细胞)传递到生殖细胞,是实现拉马克式获得性遗传的重要桥梁。
miRNA 结合 mRNA 导致降解或翻译抑制,实现转录后调控;lncRNA 可招募表观修饰酶(如 PRC2 复合物)到特定基因座,引起 H3K27me3 沉默,实现染色质水平调控;piRNA 在生殖细胞中抑制转座子,保护基因组稳定。
小 RNA(如线虫中的 piRNA/siRNA)可从体细胞传递到生殖细胞,介导回避学习记忆的跨代传递等,实现跨代表观遗传。
(4)染色质重塑。染色质重塑是指 ATP 依赖的重塑复合物(如 SWI/SNF、ISWI、CHD、INO80 家族)利用 ATP 水解能量,移动、滑动、置换或重排核小体位置和组成的过程。
它是表观遗传的“动态执行者”,它不直接添加化学标记,而是通过物理移动核小体实时调控基因表达,并帮助建立可遗传的染色质状态。在细胞分裂时,重塑状态可通过“复制”机制传递给子代细胞。有些重塑事件在发育关键窗口被“锁定”,形成稳定的表观遗传记忆。在生殖细胞重编程过程中,重塑复合物帮助清除或保留特定染色质状态,实现环境诱导的表型跨代传递。
(5)组蛋白变体的作用。常规组蛋白(H2A、H2B、H3、H4)是核小体的核心成分。组蛋白变体是这些常规组蛋白的序列略有不同的“版本”,如 H2A.X、H3.3、H2A.Z 等,它们由不同基因编码,可在特定条件下替换常规组蛋白。
不同变体使核小体更松散或更致密,从而影响基因的转录活性。例如,H3.3常与活跃转录区域相关,替换常规 H3 后使染色质更开放,促进基因表达。H2A.Z常位于启动子区域,可促进或抑制转录,参与基因激活或沉默。
组蛋白变体在 DNA 复制后可被特异性沉积酶,如 HIRA 复合物沉积 H3.3,“记住”并传递给子代细胞,甚至跨代传递,尤其在生殖细胞中。
(6)mRNA的化学修饰。如A的甲基化和C的乙酰化。可影响 mRNA 稳定性、翻译效率、剪接和定位。这是“RNA 层面”的表观遗传,补充了 DNA 和组蛋白水平,形成多层调控网络。
这些方式能被细胞乃至后代记住。关键突破是跨代表观遗传(transgenerational epigenetic inheritance):环境诱导的表观变化可通过配子传递到未暴露的后代,甚至多代。
这为拉马克学说提供了分子桥梁:环境 → 表观修饰 → 表型改变 → 可遗传。
三、重磅证据:曹晓风团队的水稻抗寒研究
2025 年 5 月 22 日,中国科学院遗传与发育生物学研究所曹晓风院士团队在 Cell 发表题为“Inheritance of acquired adaptive cold tolerance in rice through DNA methylation”的论文,首次在分子水平为拉马克“获得性遗传”提供直接证据。该研究亮点包括:

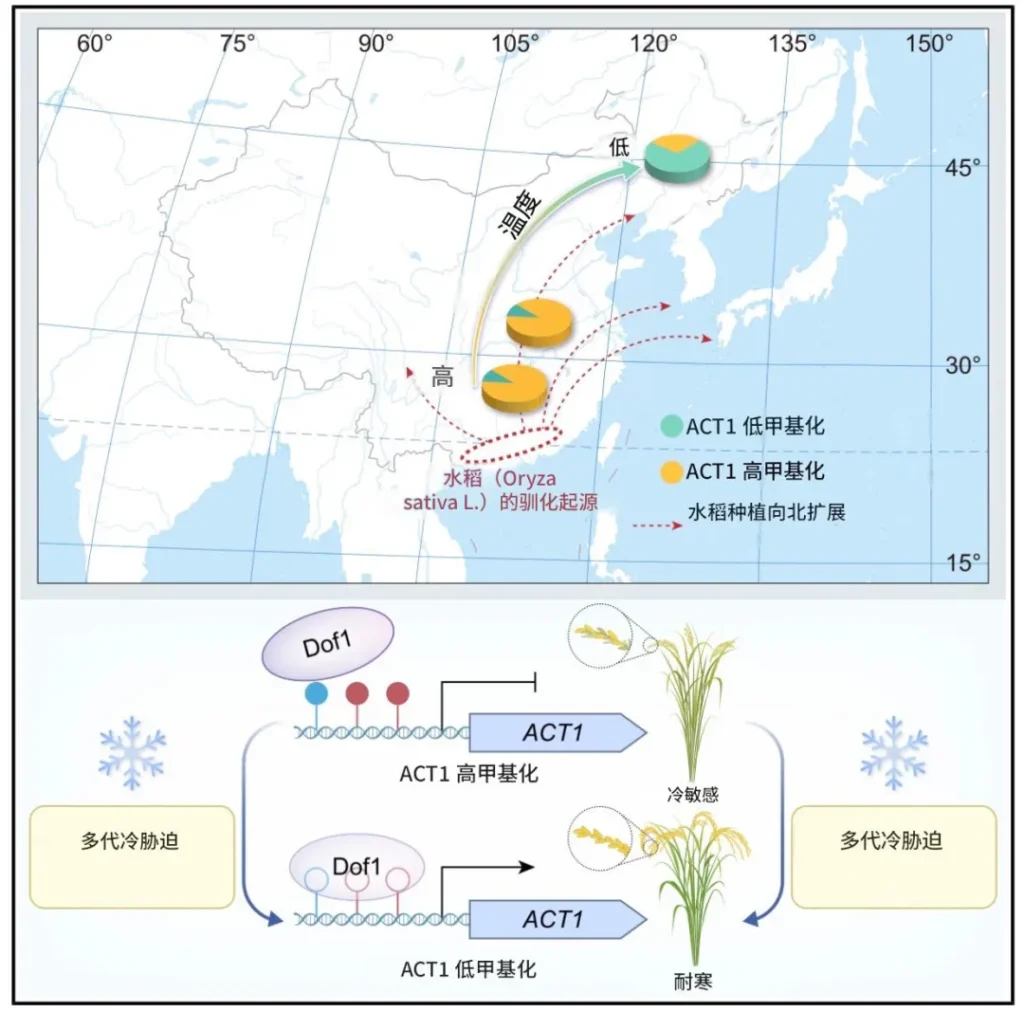
(1)建立多代连续冷胁迫筛选体系,针对水稻减数分裂期(对低温最敏感)处理。
(2)南方水稻经低温“锻炼”后,耐寒性显著提升,且显性遗传,在常温下稳定遗传至少五代。
(3)核心机制涉及阿拉伯半乳糖蛋白基因 ACT1 启动子区 DNA 去甲基化。低温抑制 DNA 甲基转移酶 MET1b 表达,导致 ACT1 表达不再受低温抑制。
(4)该表观变异在自然北移水稻中呈现“南高北低”的甲基化梯度,是适应高纬度低温的关键驯化位点。
(5)通过靶向甲基化编辑,直接验证因果关系,实现耐冷性的精准调控。
这项研究超越传统达尔文框架,为适应性进化提供新范式,并提出“逆境驯化-表观变异鉴定-精准编辑”的作物育种新思路。
这不是孤例。植物中表观遗传更常见,因为生殖细胞起源于体细胞,易受环境影响。
四、其他重要实例:涉及线虫、果蝇、小鼠、人类和原核生物
(1)秀丽隐杆线虫(Caenorhabditis elegans)。小 RNA 介导的回避学习记忆传递。线虫接触致病菌铜绿假单胞菌(Pseudomonas aeruginosa,PA14)后学会回避。这种回避行为可通过小 RNA从肠道传递到生殖细胞,稳定遗传 4 代(F1–F4),第 5 代恢复。机制涉及 piRNA 和 siRNA 放大信号,调控靶基因表达。

(2)果蝇。行为与应激适应。捕食者诱导的产卵偏好:果蝇暴露于寄生蜂(捕食者)后,会偏好在含有乙醇的食物上产卵(乙醇可保护后代免受寄生)。这种适应性行为可遗传 5 代,涉及染色质修饰和小 RNA 等表观调控。

(3)小鼠。哺乳动物中机制最清晰的模型。气味恐惧条件反射(Dias & Ressler, 2014)。雄鼠对特定气味(如乙苯乙酮)与电击配对产生恐惧。后代(F1 和 F2)对同一气味更敏感,即使未经历条件反射。机制是精子中对应嗅觉受体基因(Olfr151)发生 DNA 低甲基化,导致受体表达增加。

(4)人类。主要来自流行病学与历史队列(证据更具关联性)。荷兰饥荒(Dutch Hunger Winter, 1944–1945):孕期经历饥荒的母亲,其后代(F1)成年后代谢问题(如肥胖、糖尿病、心血管病)风险增加。F2(孙代)也有更高新生儿脂肪和健康问题风险。机制涉及 IGF2 等印记基因的 DNA 甲基化改变。该研究是人类跨代效应的经典证据。不过,人类中严格的“跨代”(F3+ 未暴露)证据较少,因为伦理限制和生命周期长,但(F1/F2)效应已较稳固,且支持“软继承”概念。

(5)原核生物。也存在表观遗传继承现象,尽管与真核生物的机制有明显差异,主要依赖DNA 甲基化和非编码RNA。原核生物是单细胞、无性繁殖,表观遗传主要体现为细胞水平的可遗传表型变异,能让子代细胞“记住”亲代经历的环境压力,实现跨代适应。
例如,一种蓝细菌( Synechocystis)氮限制应激记忆:氮缺乏诱导 DNA 甲基化(mC)模式改变。恢复正常氮供应后,许多甲基化位点仍部分保留(跨约 12 代),使后代在再次面临氮限制时生长更好。这证明了原核生物中环境诱导的跨代表观记忆。

五、 特别注意:不是简单“平反”,不是经典拉马克主义的直接回归,而是融合与扩展
表观遗传支持拉马克的部分核心,即环境诱导获得性状可遗传,但存在重要区别:
(1)稳定性。多数表观标记可逆,几代后衰减(“软继承”),难以单独驱动长期宏观进化。
(2)定向性。是反应性适应,而非拉马克想象的“内在意志”驱动。
(3)进化框架。表观机制本身是自然选择进化的产物。它补充而非取代达尔文,而是加速适应(Baldwin 效应),提供变异原料,最终受选择过滤。
(4) “硬” vs “软”。多数是暂时“软”继承;极少数可能通过表观-遗传耦合转为 DNA 序列改变的“硬”继承,但证据仍有限。
总之,表观遗传桥接了拉马克与达尔文。它提供了一种拉马克式机制,即环境直接影响可遗传性状,但置于达尔文框架下,最终受自然选择过滤,机制可进化。这推动“扩展进化综合论”(Extended Evolutionary Synthesis),承认除基因突变外,还有表观、发育、行为等多层次因素。
因此,表观遗传没有彻底推翻达尔文,而是让进化论更丰富、更动态。它提醒我们:生化世界远比 DNA 序列复杂,环境、表型与基因组在多代尺度上深度互动。
作为生化爱好者,我们正站在一个激动人心的时代——从实验室的甲基化测序,到田间的抗逆水稻,再到对自身健康的反思,表观遗传正在不断地让我们重写教科书,包括我的《生物化学原理》和《分子生物学》。

正是:
DNA 不是唯一 boss,
表观标记把记忆留。
甲基小帽子戴头上,
组蛋白尾巴舞翩跹。
非编码 RNA 传情报,
从肠道直奔生殖腺。
细菌线虫果蝇鼠,
水稻人类齐上阵。
拉马克今日笑开颜,
获得性遗传不再冤。
环境写笔记代代传,
科学进化永向前

近期评论