


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:爱基百客
很多科研新人在刚接触表观遗传领域,面对海量的文献、陌生的专业词汇以及复杂的测序技术,难免会感到一丝焦虑和千头万绪。这非常正常,每一个在这个领域深耕的学者,都是从这一步走过来的。作为在这个领域探索了许久的“老朋友”,今天我们想用最放松的方式,和你一起梳理一下表观领域最核心的一些基础概念。
01 什么是表观遗传学?
如果把我们细胞里的DNA序列(基因组)比作是一台电脑的“硬件”,那么表观遗传学就是运行在这台电脑上的“软件”。在不改变DNA序列(也就是A、T、C、G字母排列)的前提下,表观机制决定了哪些基因应该被“打开”(表达),哪些基因应该被“关闭”(沉默)。这就是为什么我们体内的神经元和心肌细胞明明拥有着完全一模一样的DNA,但它们的表型和功能却天差地别——因为它们运行的“表观软件程序”不同。
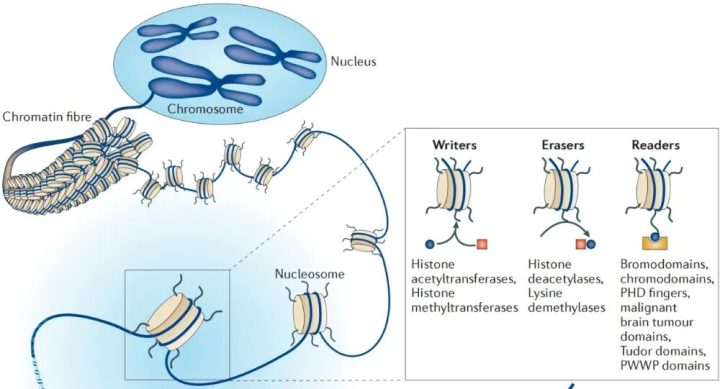
在科研语境下,表观调控系统通常由三类“打工人”组成,这也是科研新人看文献会高频遇到的概念:
- Writers(书写器):负责把表观修饰的“标签”贴到DNA或组蛋白上。
- Erasers(擦除器):负责把这些“标签”摘除,体现了表观修饰的可逆性。
- Readers(阅读器):识别这些“标签”,并招募下游的蛋白质机器来执行具体的功能(比如启动或阻止转录)。
02 表观遗传的四大表观机制
虽然表观机制种类繁多,但作为初学者,我们只需要先牢牢抓住四座大山:
第一座山:DNA甲基化——最经典、最稳定的沉默信号
DNA甲基化是最被大众熟知的表观修饰。简单来说,就是DNA的胞嘧啶(C)上被戴上了一顶“甲基”帽子(最常见的是5mC)。
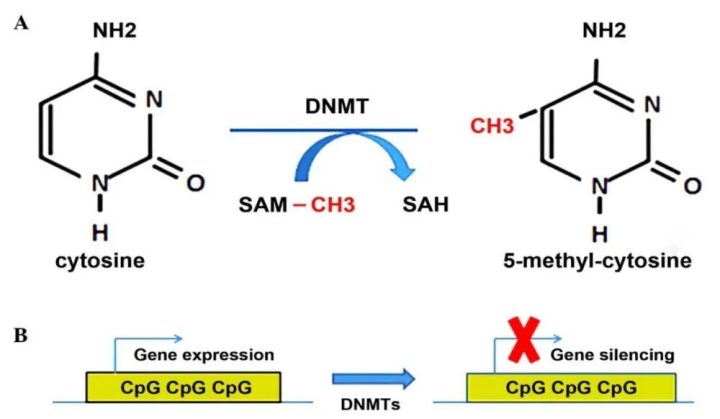
- 发生位置:在哺乳动物中,甲基化主要发生在CpG二核苷酸上(即C后面跟着G的序列)。
- 核心规律:很多基因的启动子区域富集了大量的CpG,被称为CpG岛(CpG Island)。
通常情况下,启动子区域的DNA高度甲基化,意味着这个基因被“锁死”沉默了。反之,去甲基化则通常意味着基因的激活。
这是因为启动子区域的DNA甲基化通常会抑制基因表达,因为它一方面会阻碍转录因子结合DNA,另一方面还能招募甲基化识别蛋白及其相关抑制复合物,使局部染色质变得更加致密、难以被转录机器接近;相反,去甲基化往往有助于恢复启动子的开放状态,从而促进基因激活。
在看甲基化相关的文献时,我们会经常碰到DNMT家族(负责甲基化的Writer)和TET家族(负责去甲基化的Eraser)。
第二座山:组蛋白修饰——复杂精密的“密码本”
我们的DNA并不是散落在细胞核里的,而是像线一样缠绕在“线轴”上,这个线轴就是组蛋白。DNA加组蛋白,构成了染色质。组蛋白有几条像尾巴一样的结构游离在外面,这些尾巴上氨基酸可以被加上各种化学修饰。
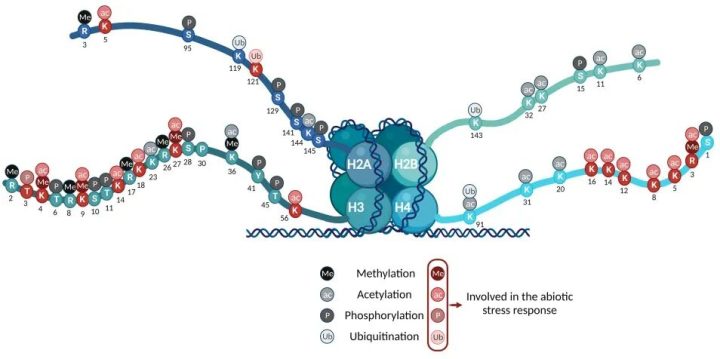
最核心的两种修饰是:
- 乙酰化(Acetylation):比如H3K27ac。乙酰化通常会让染色质结构变得松散(常染色质),方便转录机器进入,因此乙酰化几乎总是代表着基因“激活”。
- 甲基化(Methylation):组蛋白的甲基化比较“两面派”,得看具体修饰的是哪个位置或者甲基化的程度:
H3K4me1:通常位于活跃的增强子区H3K4me3:通常位于活跃基因的启动子区(激活信号)。H3K27me3和H3K9me3:通常代表基因被抑制、染色质处于致密状态(沉默信号)。
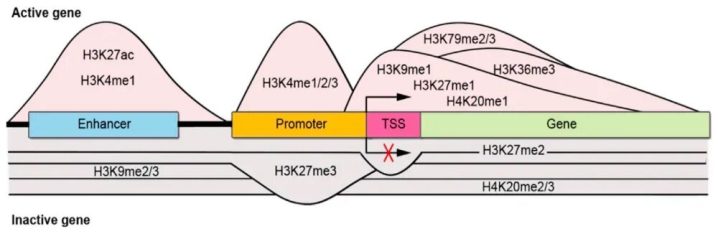
除了乙酰化和甲基化这两种常见修饰,还有一些其他修饰:巴豆酰化、磷酸化、乳酸化等,这些修饰也研究的较多的。
第三座山:染色质可及性与三维结构——宏观的空间折叠
如果我们跳出单个基因,从宏观来看,染色质在细胞核里是折叠成特定三维结构的。
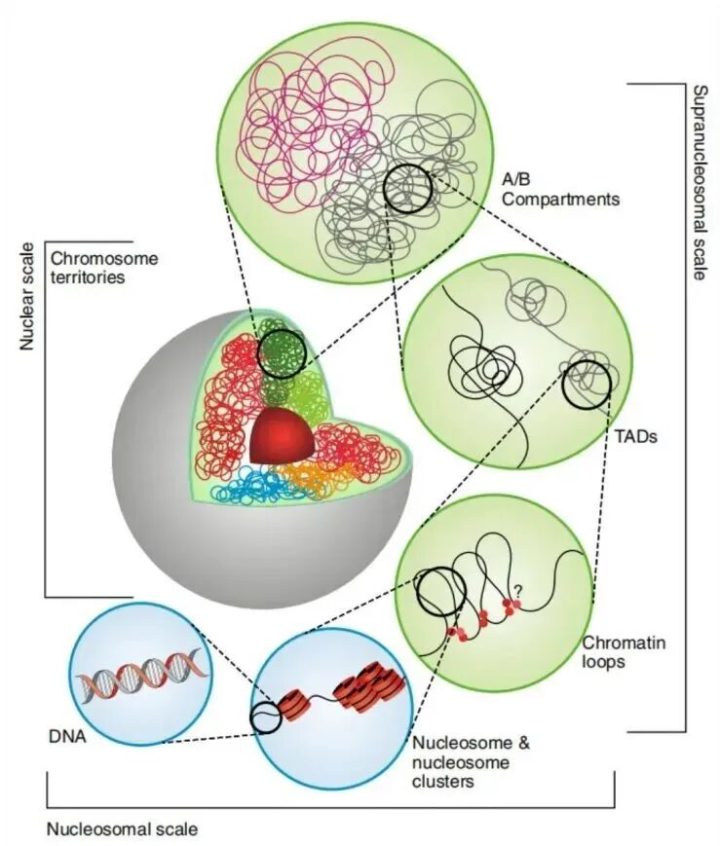
染色质三维结构
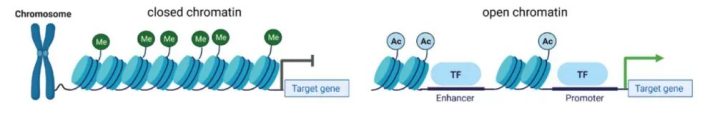
可及性(Accessibility):有些区域折叠得很紧(异染色质,关),有些区域很松散(常染色质,开)。只有“开放”的区域,转录因子才能结合上去。染色质这种开放特性,叫做染色质可及性。现在非常火的ATAC-seq技术,就是专门用来在全基因组范围内寻找这些“开放区域”的。
染色质可及性三维结构:相隔很远的增强子(Enhancer)可以通过DNA环化(Looping)在三维空间上靠近启动子(Promoter),从而调控基因。我们常用Hi-C等技术来研究这种空间上的互动。
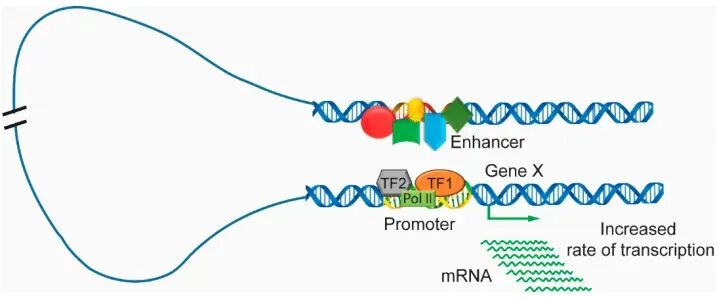 增强子-启动子环
增强子-启动子环
第四座大山:染色质重塑——基因组的“硬核搬运工”
如果说DNA甲基化和组蛋白修饰是贴在基因组上的“指令标签”,那么染色质重塑(Chromatin Remodeling)就是真正去执行物理搬运的“工人”。
前面我们提到,DNA是缠绕在组蛋白这个“线轴”上的(形成核小体)。当某个基因需要被强烈表达时,转录机器(转录因子/辅因子、RNA聚合酶等)需要一块宽敞的场地来工作。这时候,光有“这里需要开放”的标签是不够的,必须有力量把挡路的“线轴(组蛋白)”移走。染色质重塑复合物就是干这个苦力活的。它们像是一台台微型的分子马达,通过消耗能量(ATP),在染色质上执行以下几种操作:
(1)逐出(Ejection):直接把碍事的组蛋白给踢走,腾出大片空间。
(2)滑动(Sliding):把核小体沿着DNA往前推一推,把原本藏在底下的DNA序列暴露出来。
(3)替换(Exchange):替换上特殊的组蛋白变体。
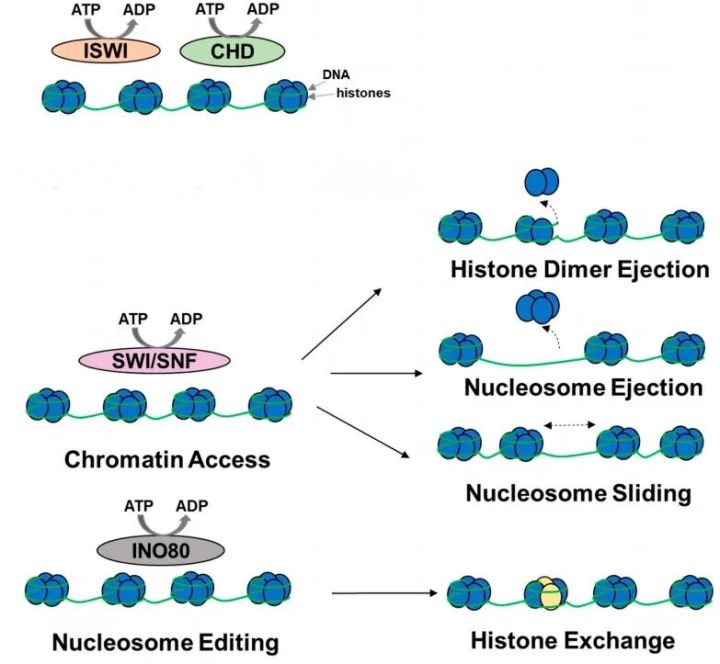
在阅读文献时,你一定会频繁遇到染色质重塑复合物:SWI/SNF(也叫BAF)复合物、ISWI、CHD等家族。尤其是SWI/SNF复合物,近年来在肿瘤研究领域简直红得发紫,因为科学家们发现超过20%的人类癌症中都存在这类“搬运工”的基因突变。
03 其他表观概念
除了上面提到的机制,我们继续补充一些文献中常见的概念:
01 表观重编程 (Epigenetic Reprogramming)
如果说表观修饰是给系统装了各种软件,那表观重编程就是“格式化硬盘,恢复出厂设置”。在生命的早期(精卵结合后形成受精卵的过程中),几乎所有的DNA甲基化表观标记都会被大规模擦除,让细胞回到最原始、最具潜能的“白纸”状态,然后再根据发育需要重新写上新的表观密码。
02 表观记忆 (Epigenetic Memory)
我们的大脑有记忆,细胞其实也有。表观记忆是指细胞在经历过某种环境刺激(比如短暂的饥饿、炎症、或者是某种药物处理)后,哪怕这个刺激已经消失了,细胞依然能通过表观修饰把这段经历“记”下来,甚至传给它的子代细胞。
03 二价染色质 (Bivalent Chromatin)
这是一个极其高频且非常有意思的专业概念,尤其在干细胞和发育生物学领域。我们前面提到,H3K4me3代表“激活”(油门),H3K27me3代表“沉默”(刹车)。通常它们是不会同时出现在同一个基因上的。但在干细胞里,很多关键的发育基因启动子上,居然同时带有这两种修饰! 为什么要这样?因为干细胞处于一种“蓄势待发(Poised)”的状态。带着“二价修饰”的基因处于微弱沉默或极低表达状态,但一旦细胞接收到分化信号,它只需要松开刹车(去掉H3K27me3),基因就能瞬间全速启动;或者松开油门(去掉H3K4me3),基因就会关闭。这展现了表观调控极高的灵活性。
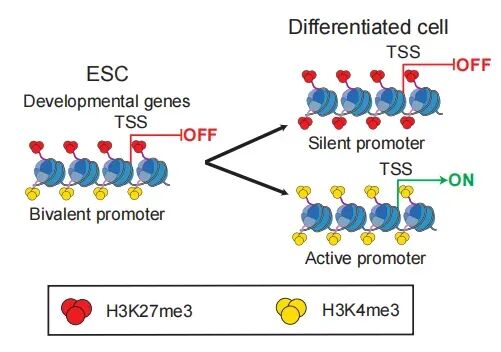
二价启动子在胚胎干细胞和分化细胞中的不同状态及其转变过程
04 表观时钟 (Epigenetic Clock)
这是近十年衰老研究领域最火爆的概念之一。随着我们年龄的增长,DNA序列本身几乎不怎么变,但是DNA上的甲基化模式却会发生极其规律的改变(有些地方甲基化越来越高,有些地方越来越低)。科学家利用机器学习算法,只要检测你血液里某些特定CpG位点的甲基化水平,就能极其精准地推算出你的“生物学年龄”。
经常看文献会看到“Horvath Clock(霍瓦斯时钟)”。现在的研究不仅用它来测年龄,还在研究各种因素(如熬夜、抽烟、运动、某种抗衰老药物)是会让你的表观时钟“加速转动”,还是“逆转倒流”。



近期评论