

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:师者解忧馆
那些曾被我们贴上“简单”、“原始”标签的原核生物,其内部世界的精巧与复杂,正逐渐浮出水面,挑战着教科书里的经典定义。
一、拟核DNA:并非完全“裸奔”
传统教材告诉我们,原核细胞的拟核由裸露的环形DNA构成,没有蛋白质包裹。这似乎成了它与真核细胞染色体最直观的区别。
然而,真相更为微妙。研究发现,细菌中存在一类被称为 “拟核结合蛋白” 的蛋白质。它们虽然不是真核细胞中的组蛋白,但功能上堪称 “简化版组蛋白”。这些NAPs能与DNA非特异性地结合,压缩染色体结构,并参与基因的转录调控。它们的存在意味着,原核生物的DNA并非在细胞质中完全“裸奔”,而是有自己独特的“管理团队”。
尽管如此,这种结合远不如真核细胞中DNA与组蛋白的紧密结合那样稳定和高度组织化。因此,在本质上,我们仍可以说拟核DNA是相对裸露的,但必须认识到其背后存在精细的调控机制。
二、多细胞形态:原核世界的“特例”
“原核生物都是单细胞的”,这是另一个根深蒂固的观念。的确,绝大多数细菌、古菌以单细胞形态存在,形态无非球、杆、螺旋几种。但生命总是充满例外。科学家发现了 多细胞趋磁原核生物。它们由多个细胞聚集而成,形成一个球状或卵球状的“多细胞”集合体。
更神奇的是,这个集合体作为一个整体,能够沿着地球磁力线定位和运动,而其中的单个细胞并不具备此能力。这就像一艘微型的“磁力潜艇”,展现了原核生物在特定环境下演化出的简单协作形态。
三、隐秘骨架:支撑细胞的内部框架
细胞骨架曾被认为是真核生物的“专利”,是维持细胞形态、实现胞内运输和分裂的基础。但原核生物真的只是一团“没有骨架的软泥”吗?
当然不是。在原核细胞内,科学家发现了功能与结构上与真核细胞骨架惊人相似的蛋白质。
FtsZ蛋白,在结构和聚合方式上类似微管蛋白,它在细胞分裂部位形成“Z环”,主导细菌的一分为二。
MreB蛋白,则像肌动蛋白丝,在杆状细菌中形成螺旋丝状结构,是维持细胞“身材”的关键。
这些发现彻底打破了“原核无骨架”的旧观念,揭示了从细菌到人类,维持细胞形态的基本原理有着深远的同源性。
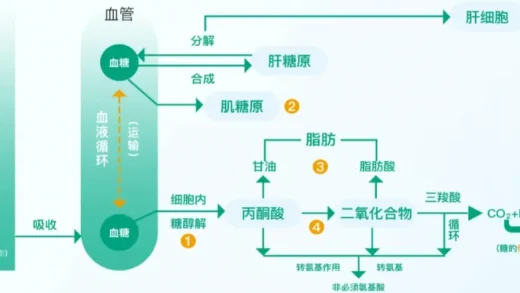
四、呼吸场所:没有线粒体,怎么办?
真核细胞的有氧呼吸“三部曲”清晰明了:细胞质基质→线粒体基质→线粒体内膜。那么,没有线粒体的原核生物如何进行有氧呼吸?
答案是:它们把整套酶系统“安装”在了不同的地方。
第一阶段,在细胞质基质中分解葡萄糖,这与真核细胞相同。第二阶段,催化三羧酸循环的酶也分布在细胞质基质中,而非线粒体基质。最关键的第三阶段,即产生大量能量的电子传递链,其相关的酶系则镶嵌在细胞膜上。原核生物的细胞膜,因此承担了线粒体内膜的功能。
这一对比鲜明地体现了生命的适应性与统一性:实现相同功能的生化途径,可以通过不同的细胞结构来完成。
五、等位基因与基因型
在遗传学中,“等位基因”通常指位于同源染色体相同位置、控制相对性状的基因。显然,这套基于二倍体有性生殖的定义,对通常为单倍体、无典型有性生殖的原核生物不太适用。
现代遗传学对等位基因的定义已拓宽为:同一基因的不同形式。因此,一个细菌基因发生突变后产生的不同变异体,也可以被视为等位基因。
至于基因型,如果狭义地理解为“等位基因的组合方式”,则原核生物确实难以符合。但若广义地理解为 “个体全部的基因组成”,那么所有生物,包括原核生物,都拥有自己的基因型。
不过在实践中,对于原核生物,科学家更常使用 “基因组” 这一术语来描述其遗传组成,这避免了概念上的纠结。
六、细胞壁:坚固堡垒的化学密码
原核生物的细胞壁是其标志性结构,主要由肽聚糖构成。肽聚糖是一个网状大分子,由聚糖链(由N-乙酰葡萄糖胺和N-乙酰胞壁酸交替连接)和肽链(四肽尾和肽桥)交织而成。
这解释了为什么蛋白酶不能彻底破坏细胞壁:蛋白酶只能水解肽键,而对连接聚糖链的 β-1,4-糖苷键 无能为力。真正能有效瓦解这堵“墙”的是溶菌酶,它专门攻击这种糖苷键。
革兰氏阳性菌和阴性菌细胞壁的复杂差异,更是展现了原核生物在细胞外围防御结构上的多样化策略。
随着研究的深入,原核生物看似简单的面纱被层层揭开,展现出一个在微观尺度上高度复杂、精妙且多样化的世界。这些发现不仅修正了我们的教科书知识,更深刻地揭示了生命演化历程中,解决相同生存问题的不同方案。
从“简单原始”到“精巧复杂”,我们对原核生物的认知变迁本身,就是一部生动的科学探索史。它提醒我们,科学知识永远是动态发展的,对任何生命形式的敬畏与好奇,才是推动认知前进的根本动力。
来源网址:一直误解原核生物?远比你想象的更“高级”

近期评论