

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:生物奇迹biomiracle
mRNA 的翻译方向从 5’端到 3’端是由其分子结构、核糖体工作机制及生物学功能共同决定的,这种方向性对蛋白质合成的准确性和效率至关重要,以下从多个维度详细解析其原因。
核糖体的结构与移动方向
1、核糖体的读取机制
核糖体是翻译的核心场所,由大小亚基组成。在翻译起始阶段,小亚基先结合 mRNA 的 5’端(通常依赖 5’帽子结构的识别),然后沿 mRNA 向 3’端移动,逐个读取密码子。这种移动方向是由核糖体的结构决定的 —— 其内部的 RNA 结合通道和肽基转移酶活性位点的排列,使得 mRNA 只能以 5’→3’的方向穿过核糖体,确保密码子按顺序被识别。
2、tRNA 的进位顺序
翻译过程中,tRNA 携带氨基酸依次进入核糖体的 A 位点(氨酰位)、P 位点(肽酰位)和 E 位点(退出位)。核糖体每读取一个密码子,就会推动 mRNA 移动三个核苷酸(一个密码子的长度),这个过程严格遵循 5’→3’的方向,以保证 tRNA 按正确顺序匹配密码子。
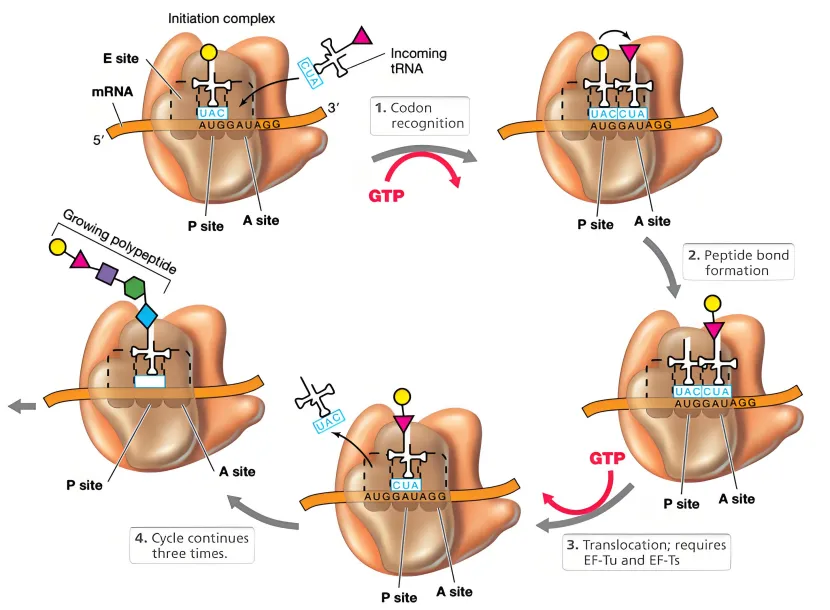
氨基酸链的合成方向(N 端→C 端)
蛋白质的基本结构是氨基酸通过肽键连接形成的多肽链,其合成方向是从 N 端(氨基端)到 C 端(羧基端)。这一方向与 mRNA 的 5’→3’翻译方向直接对应:
第一个进入核糖体的 tRNA 携带甲硫氨酸(起始密码子 AUG 对应的氨基酸),作为多肽链的 N 端起点;
后续 tRNA 携带的氨基酸通过肽基转移酶的作用,依次连接到前一个氨基酸的羧基上,形成肽链的 C 端延伸。
因此,mRNA 的翻译方向(5’→3’)与多肽链的合成方向(N 端→C 端)是同步的,这种一致性确保了蛋白质一级结构的正确构建。
mRNA 的结构特征与起始信号
1、5’端帽子结构的作用
真核生物 mRNA 的 5’端通常有一个甲基化的帽子结构(m7GpppN),它不仅能保护 mRNA 免受核酸酶降解,还能被起始因子(如 eIF4E)识别,引导核糖体结合到 mRNA 的 5’端,从而确定翻译的起始位置。由于起始信号位于 5’端附近,核糖体自然从该端开始向 3’端移动。
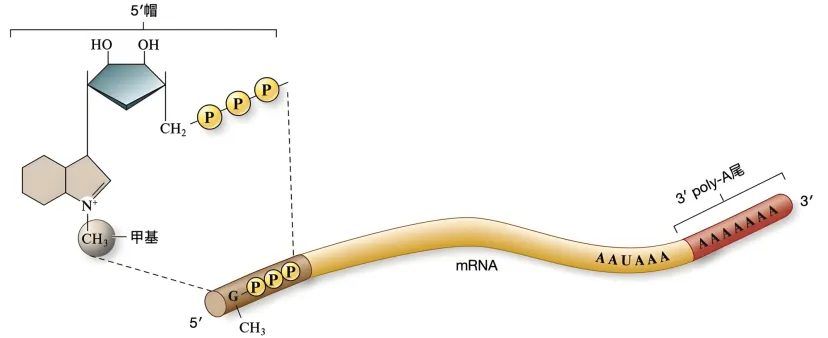
2、起始密码子的位置
翻译起始需要识别起始密码子(通常为 AUG),而 AUG 在 mRNA 上的位置靠近 5’端。核糖体从小亚基结合 5’端开始,沿 mRNA 扫描直至找到 AUG,随后大亚基结合并启动翻译。这种扫描机制决定了翻译必须从 5’端向 3’端进行。
转录与翻译的偶联(原核生物)
在原核生物中,转录和翻译可以同时进行——RNA 聚合酶沿 DNA 模板链的 3’→5’方向移动,以NTP为原料,按5’→3’方向合成 mRNA,而核糖体则紧随转录复合物,在 mRNA 尚未完全合成时就从其 5’端开始翻译。这种偶联机制要求翻译方向与转录方向一致(均为 5’→3’),以提高基因表达的效率。
进化与功能优势
1、错误校正机制
从 5’→3’翻译时,若核糖体遇到异常密码子或 mRNA 损伤,可及时终止翻译并释放错误产物,避免浪费能量合成无效蛋白质。这种方向性使得质量控制机制能按顺序监控翻译过程。
2、mRNA 稳定性与降解
3’端的 poly-A 尾巴能延长 mRNA 的半衰期,而翻译从 5’端开始可优先利用完整的 mRNA 模板。当 mRNA 逐渐被降解时(通常从 5’端开始),核糖体仍能优先翻译出完整的蛋白质序列,提高翻译效率。
实验证据支持
通过放射性同位素标记实验(如用 3H 标记氨基酸),可观察到新合成的多肽链中,N 端先被标记,C 端后被标记,直接证明了翻译过程中核糖体沿 mRNA 从 5’→3’移动,多肽链从 N 端向 C 端延伸。
总结
mRNA 的 5’→3’翻译方向是分子结构、功能需求与进化选择的共同结果:核糖体的移动机制、氨基酸链的合成方向、mRNA 的起始信号定位,以及转录 – 翻译偶联等因素,共同确保了这一方向性的高度保守性。这种机制不仅保证了蛋白质合成的准确性,还优化了基因表达的效率,是生命系统精密调控的典型体现。



近期评论