

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号: 卢镇岳斋聊生物学
摘要:本文深入综述了植物与真菌、原核生物及病毒之间相互作用的分子机制。在植物与真菌的相互作用方面,详细探讨了共生及致病过程中的分子信号与机制;植物与原核生物的相互作用涵盖了根际微生物促进植物生长、植物病原菌致病以及内共生细菌与植物代谢协作的分子策略;植物与病毒的相互作用则包括病毒入侵、植物抗病毒免疫反应及两者共进化的分子过程。这些研究有助于深入理解植物生态系统中的生物关系,为农业、生态等领域提供重要理论基础。
关键词:植物;真菌;原核生物;病毒;分子机制
一、植物与真菌的相互作用
(一)共生关系中的分子信号
1. 植物信号分子的分泌与作用
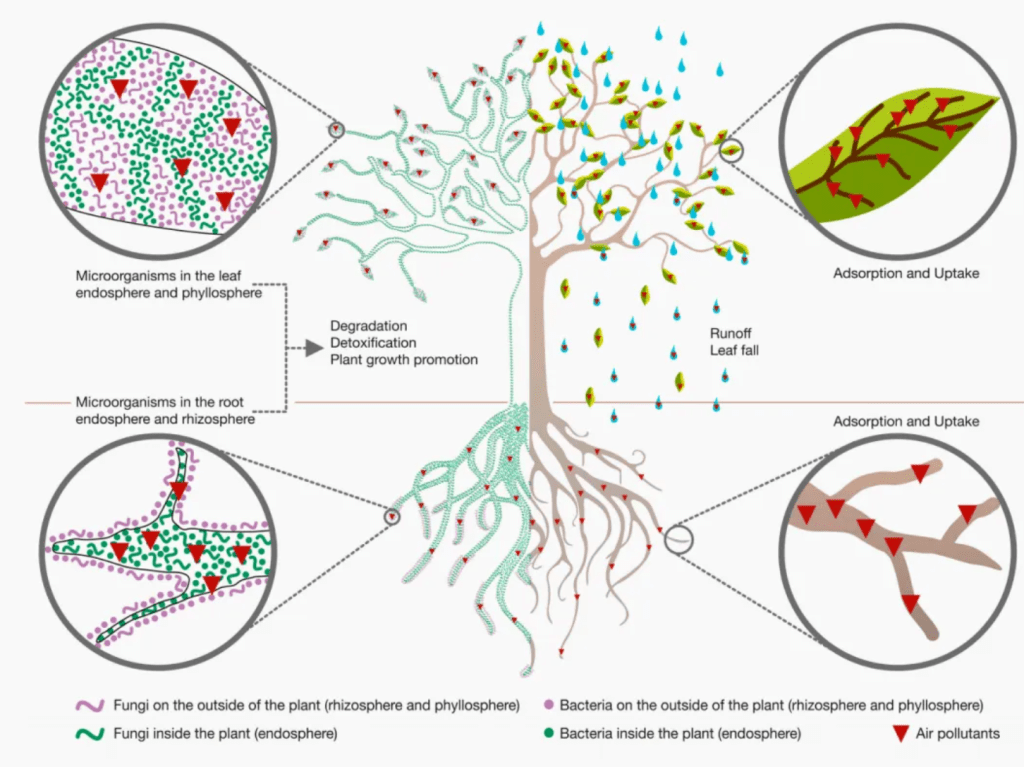
植物根系在与真菌建立共生关系时,会分泌独脚金内酯等信号分子。独脚金内酯作为一类重要的植物激素,能够在低浓度下被丛枝菌根真菌(AMF)感知。其作用机制在于,独脚金内酯可诱导真菌菌丝的分枝和生长方向的改变,促使真菌向植物根系靠近,为后续的共生关系建立奠定基础。在分子层面,独脚金内酯与真菌细胞膜上的受体结合,激活下游信号通路,引发真菌细胞内一系列生理生化反应,如钙离子浓度的变化等,这些变化进一步调控真菌的基因表达,促进与共生相关的基因转录,增强真菌对植物根系环境的适应性和侵染能力。
2. 真菌信号的反馈与共生建立
真菌在接收到植物的信号后,会产生 Myc 因子等信号分子作为回应。Myc 因子能够被植物细胞表面的受体识别,进而激活植物细胞内的共生信号通路。在这个过程中,植物细胞内会发生钙离子振荡,这是共生信号传递的关键事件之一。钙离子振荡会激活钙调蛋白依赖性蛋白激酶等一系列蛋白激酶,这些激酶通过磷酸化作用调控下游基因的表达。例如,它们会促进与根瘤菌共生相关基因(如 SymRK 基因)的表达,SymRK 蛋白作为植物共生受体激酶,在植物与真菌的信号传递和共生关系建立中起到核心枢纽作用,其通过磷酸化级联反应,调节植物根系细胞的分化和代谢,为真菌的侵染和定殖创造有利条件,最终实现植物与真菌的紧密共生。
(二)致病过程中的分子机制
1. 致病真菌效应蛋白的作用机制
植物致病真菌分泌的效应蛋白在致病过程中发挥着关键作用。这些效应蛋白通常具有靶向植物细胞内特定蛋白或信号通路的能力。例如,某些效应蛋白能够与植物细胞内的免疫受体蛋白相互作用,干扰其正常功能,从而抑制植物的 PAMP 触发的免疫反应(PTI)。具体而言,效应蛋白可能通过模拟植物自身蛋白的结构或功能,与免疫受体竞争结合底物,或者直接修饰免疫受体,使其无法正常激活下游的防御信号转导,导致植物细胞内防御基因的表达受阻,植物的免疫系统无法有效启动,使得真菌能够顺利侵染植物组织,在植物体内定殖和繁殖,引发植物病害症状。
2. 细胞壁降解酶的作用与影响
真菌分泌的细胞壁降解酶是其突破植物细胞壁防线的重要武器。纤维素酶能够特异性地分解植物细胞壁中的纤维素成分,果胶酶则作用于果胶物质。在分子水平上,这些酶通过水解细胞壁多糖的糖苷键,破坏细胞壁的结构完整性。例如,纤维素酶的活性位点能够与纤维素分子紧密结合,催化水解反应,使纤维素长链断裂为较短的片段。细胞壁的破坏不仅为真菌菌丝的侵入提供了物理通道,还会引发植物细胞的一系列应激反应。植物细胞会感知细胞壁的损伤信号,启动修复机制,但在致病真菌的持续攻击下,植物细胞往往难以有效应对,导致真菌能够在植物组织内蔓延,进一步摄取植物细胞的营养物质,严重影响植物的生长和健康,造成农业生产上的损失。
二、植物与原核生物的相互作用
(一)根际微生物促进植物生长的分子机制
1. 植物激素的合成与调控
根际促生细菌(PGPR)能够合成多种植物激素,其中生长素的合成途径较为复杂。在色氨酸依赖途径中,PGPR 利用色氨酸为前体,通过一系列酶促反应合成生长素。例如,色氨酸在色氨酸转氨酶等酶的作用下,逐步转化为吲哚 – 3 – 丙酮酸,再经过脱羧等反应生成生长素。合成的生长素通过主动运输或扩散等方式进入植物细胞。在植物细胞内,生长素与生长素受体结合,激活受体蛋白的激酶活性,引发下游信号转导。这会导致植物细胞的质子 – ATP 酶基因表达上调,使细胞外环境酸化,细胞壁松弛,从而促进植物细胞的伸长和分裂,特别是在植物根系细胞中,生长素的作用使得根系生长更加旺盛,增加了植物根系的表面积和吸收能力,有助于植物更好地吸收水分和养分,促进植物整体的生长发育。
2. 铁载体的功能与作用机制
PGPR 产生的铁载体如荧光假单胞菌分泌的嗜铁素,具有高度特异性的铁离子结合能力。嗜铁素分子结构中含有多个能够与铁离子配位结合的官能团,其能够在土壤环境中与铁离子形成稳定的复合物。在低铁条件下,PGPR 会大量合成并分泌嗜铁素,通过主动运输将铁 – 嗜铁素复合物摄取到细菌细胞内,满足自身对铁元素的需求。同时,这一过程改变了根际土壤中铁的化学形态和有效性,使得原本难以被植物吸收的铁变得更容易被植物利用。植物细胞表面存在能够识别铁 – 嗜铁素复合物的转运蛋白,这些转运蛋白将铁 – 嗜铁素复合物转运到植物细胞内,经过细胞内的代谢过程,铁离子被释放并参与植物的光合作用、呼吸作用等重要生理过程,如铁是植物细胞色素和铁硫蛋白等的重要组成成分,对植物的能量代谢和电子传递过程至关重要,从而促进植物的生长和健康。
(二)植物病原菌致病的分子策略
1. Ⅲ 型分泌系统与效应蛋白的作用
植物致病细菌的 Ⅲ 型分泌系统是其致病的关键装置。该系统由多个蛋白组成,形成一个类似注射器的结构,能够将效应蛋白直接注入植物细胞内。例如,青枯雷尔氏菌的 Ⅲ 型分泌系统识别并转运效应蛋白进入植物细胞后,这些效应蛋白会干扰植物细胞的正常代谢和信号转导过程。一些效应蛋白能够与植物细胞内的 MAPK 信号通路中的关键激酶相互作用,抑制激酶的活性,从而阻断植物细胞对病原菌侵染的防御信号传递。这使得植物细胞无法正常激活防御相关基因的表达,如植保素合成基因、病程相关蛋白基因等,导致植物的免疫系统瘫痪,无法有效抵御病原菌的入侵,使得细菌在植物体内大量繁殖,引发植物病害症状,如叶片枯萎、植株生长受阻等,严重影响植物的生长和存活,对农业生产造成巨大威胁。
2. 毒素与其他致病因子的分子机制
除了效应蛋白,一些植物病原菌还会产生毒素等致病因子。例如,镰刀菌产生的镰刀菌毒素,其作用机制包括抑制植物细胞的蛋白质合成过程。在分子水平上,镰刀菌毒素能够与植物细胞的核糖体结合,干扰核糖体的功能,阻止 mRNA 与核糖体的正常结合和翻译过程,导致植物细胞内蛋白质合成受阻,影响植物细胞的正常代谢和生理功能。病原菌产生的其他致病因子如胞外多糖等,能够帮助病原菌在植物组织内定殖和扩散。胞外多糖可以形成一层黏性物质,保护病原菌免受植物防御机制的攻击,同时也有助于病原菌与植物细胞表面的黏附,促进病原菌在植物组织中的侵染和传播,进一步加重植物病害的程度。
(三)内共生细菌与植物的代谢协作分子机制
1. 根瘤菌固氮的分子过程
根瘤菌与豆科植物共生过程中,固氮作用是其重要的功能。根瘤菌的固氮酶基因在共生关系建立后被激活表达。固氮酶是由多个亚基组成的复杂酶系统,其催化氮气还原为氨的过程需要消耗大量的能量和还原力。在根瘤内的微环境中,根瘤菌通过呼吸作用产生的能量和电子传递链提供的还原力,驱动固氮酶将氮气逐步还原为氨。生成的氨通过根瘤菌的氨转运蛋白运输到植物细胞内,在植物细胞内,氨首先被转化为谷氨酸等氨基酸,然后进一步参与植物体内蛋白质、核酸等含氮生物大分子的合成。这一过程为植物提供了稳定的氮源,减少了植物对土壤中氮肥的依赖,在生态农业中具有重要意义,有助于提高土壤肥力和减少环境污染。
2. 内生细菌与植物次生代谢的关联
某些内生细菌能够参与植物的次生代谢过程。例如,一些内生细菌能够诱导植物合成植保素等次生代谢产物。在分子层面,内生细菌可能通过分泌信号分子或与植物细胞表面受体相互作用,激活植物细胞内的次生代谢相关信号通路。这会导致植物细胞内次生代谢基因的表达上调,如苯丙烷代谢途径中的关键基因。苯丙烷代谢途径被激活后,会合成一系列次生代谢产物,其中植保素具有抗菌、抗虫等生物活性,能够增强植物对病虫害的抵抗能力。此外,内生细菌还可能影响植物的品质和风味物质的形成,如某些内生细菌能够促进植物合成挥发性有机化合物,这些化合物不仅影响植物的风味,还可能在植物与其他生物的相互作用中发挥信号传递作用,对植物的生态适应性和经济价值产生重要影响。
三、植物与病毒的相互作用
(一)病毒入侵植物的分子过程
1. 病毒与植物细胞受体的识别与结合
植物病毒入侵植物的第一步是其衣壳蛋白与植物细胞表面受体的识别和结合。不同的植物病毒具有不同的衣壳蛋白结构,能够特异性地与植物细胞表面的相应受体结合。例如,烟草花叶病毒的衣壳蛋白能够与烟草细胞表面的特定蛋白受体相互作用。这种结合具有高度的特异性和亲和力,其分子机制涉及到衣壳蛋白和受体蛋白之间的静电相互作用、氢键和疏水相互作用等。一旦结合发生,会触发植物细胞的内吞作用或膜融合过程,使病毒粒子进入植物细胞内。在一些情况下,病毒与受体的结合还会引发植物细胞表面的信号转导,激活或抑制某些细胞内的信号通路,但目前对于这些信号通路的具体作用和调控机制仍不完全清楚,需要进一步深入研究。
2. 病毒基因组的复制与表达
进入植物细胞后的病毒,利用植物细胞的转录和翻译系统进行自身基因组的复制和蛋白质合成。病毒的 RNA 依赖的 RNA 聚合酶(RdRp)在这一过程中起到关键作用。RdRp 能够以病毒 RNA 为模板,按照碱基互补配对原则合成互补 RNA(cRNA),然后以 cRNA 为模板进一步复制出大量的病毒基因组 RNA。在病毒蛋白质合成过程中,病毒 RNA 会与植物细胞的核糖体结合,利用植物细胞的氨基酸和能量代谢系统合成病毒的结构蛋白和非结构蛋白。病毒的非结构蛋白通常参与病毒基因组的复制和调控过程,而结构蛋白则用于组装新的病毒粒子。例如,病毒的衣壳蛋白在细胞质中合成后,会与病毒基因组 RNA 相互作用,在特定的细胞区域内进行病毒粒子的组装,为病毒的传播和进一步侵染做准备。
(二)植物抗病毒的免疫反应分子机制
1. RNA 沉默机制的作用过程
植物的 RNA 沉默机制是其抗病毒的重要防线。当植物细胞识别到病毒的双链 RNA(dsRNA)后,会启动 RNA 沉默途径。首先,Dicer 样酶会识别并切割病毒 dsRNA,将其切割成 21 – 24 核苷酸长度的小干扰 RNA(siRNA)。这些 siRNA 具有与病毒 RNA 互补的序列,它们会与 Argonaute 蛋白结合形成 RNA 诱导沉默复合体(RISC)。RISC 中的 Argonaute 蛋白具有核酸内切酶活性,能够识别并降解与 siRNA 互补的病毒 RNA。在这个过程中,植物细胞内还存在一些辅助蛋白参与 RISC 的组装和功能调控,如双链 RNA 结合蛋白等。RNA 沉默机制能够有效地抑制病毒的复制和传播,保护植物免受病毒的侵害,但病毒也会进化出一些机制来逃避植物的 RNA 沉默反应,如编码抑制植物 RNA 沉默的蛋白,这导致了植物与病毒之间在 RNA 沉默层面的军备竞赛。
2. 抗性基因介导的免疫反应
植物的抗性基因(R 基因)在抗病毒过程中发挥关键作用。当植物携带与病毒无毒基因相对应的 R 基因时,病毒侵染会触发植物的效应子触发免疫(ETI)反应。R 基因编码的蛋白通常具有核苷酸结合位点(NBS)和富含亮氨酸重复序列(LRR)等结构域。这些结构域使得 R 基因编码的蛋白能够识别病毒的效应蛋白。例如,当病毒的效应蛋白进入植物细胞后,R 基因编码的蛋白通过其 LRR 结构域与病毒效应蛋白相互作用,这种相互作用会激活植物细胞内的防御信号通路,如激活植物的 MAPK 信号通路和钙离子信号通路等。这些信号通路的激活会引发植物的超敏反应,使受侵染的植物细胞程序性死亡,形成局部坏死斑,限制病毒的进一步扩散。同时,激活的信号通路还会诱导植物合成并积累病程相关蛋白、植保素等防御物质,增强植物对病毒的抗性和对后续侵染的免疫能力。
(三)病毒与宿主的共进化分子机制
1. 病毒的进化适应策略
在长期的相互作用中,植物病毒为了逃避植物的免疫识别和防御机制,不断进化其基因组和蛋白序列。病毒的效应蛋白是其与植物免疫系统对抗的关键武器,病毒会通过点突变、基因重组等方式改变效应蛋白的结构和功能。例如,一些病毒的效应蛋白通过突变获得了抑制植物 RNA 沉默机制或干扰植物抗性基因介导的免疫反应的能力。病毒还会改变其传播方式和宿主范围,以适应不同的生态环境和植物宿主。某些病毒可能通过获得新的基因或改变基因表达模式,使其能够借助新的媒介昆虫进行传播,或者能够感染原本具有抗性的植物品种,从而增加其在生态系统中的生存和传播能力。
2. 植物的防御进化与适应
植物在与病毒的共进化过程中,也不断进化其防御机制。一方面,植物通过基因突变和基因复制等方式产生新的抗性基因或改变现有抗性基因的功能。例如,植物可能会产生具有更高亲和力和特异性的 R 基因变体,能够识别更广泛的病毒效应蛋白。另一方面,植物会优化其免疫信号通路和防御代谢途径。植物可能会增强 RNA 沉默机制的效率,增加 Dicer 样酶的活性或提高 RISC 的稳定性,使其能够更有效地降解病毒 RNA。同时,植物在受到病毒侵染后,会调整其生长发育策略,如延迟开花、减少营养生长等,以减少病毒对其繁殖和生存的影响,提高自身在病毒侵染压力下的生存能力和适应性,维持植物种群在生态系统中的稳定。
四、总结与展望
植物与真菌、原核生物和病毒之间的相互作用通过复杂多样的分子机制紧密相连,这些机制深刻影响着植物的生长、健康和生态系统的平衡。在未来的研究中,随着基因编辑技术如 CRISPR-Cas 系统的不断发展和完善,有望更加精准地解析这些相互作用中的关键基因和分子过程。
单细胞测序技术的应用将使我们能够深入了解单个细胞在植物 – 微生物 / 病毒相互作用中的反应和功能差异。在农业生产领域,基于对这些分子机制的深入理解,可以开发出更具针对性和高效性的生物防治策略,如利用基因编辑技术培育具有更强抗病虫能力的植物品种,或者筛选和应用更有效的根际促生微生物。在生态保护方面,有助于揭示生态系统中生物间相互关系的分子基础,为生态修复和生态系统功能维持提供理论指导。
然而,目前仍存在许多未知领域,例如微生物群落中不同物种间跨界分子信号网络的精细结构和动态变化、病毒与宿主在复杂生态环境下共进化的详细分子轨迹等,需要进一步整合多学科技术和方法进行深入探究,以全面揭示植物与微生物及病毒相互作用的奥秘,为生物科学的发展和应用提供更坚实的基础。
来源网址:看不见的植物关系试题情境满满了


近期评论