


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
李晓燕 孙鹏飞 王艳 吴志强 王健
摘要:结合表观遗传景观图的核心寓意,从现象层面解析高中生物学教材中植物组织培养、核移植和iPS细胞技术中所涉及的细胞全能性的问题。以多因素调控为核心解析细胞全能性表达的分子机制,为高中生物学教师教学和学生深度理解细胞全能性提供参考。
高中生物学中不同版本教材对细胞全能性概念的描述存在细微差别,但都强调细胞全能性本质上是一种潜能,并不一定能表达出来。理论上,某一细胞具有发育成完整个体所需要的全部遗传信息,则该细胞具有全能性。细胞全能性虽依赖全套的遗传信息,但不是一种即时表现的能力,也不是在任何时刻都能表达的普遍现象,而是特定的细胞类型在特定条件和特定的发育阶段才有可能被激活的潜能。细胞全能性是植物组织培养与核移植技术的理论基础。在教学过程中,教师与学生常常难以将细胞全能性的相关问题与表观遗传建立起联系,无法深入理解细胞全能性的多因素调控。本文结合科研资料,对高中生物学中有关细胞全能性的相关问题进行解析,为教学提供参考。
1 表观遗传景观图的提出
受精卵如何发育成复杂的生物个体?受精卵分裂、分化过程中,为什么有些细胞形成心肌细胞、有些细胞形成骨骼肌细胞?1957年,英国发育生物学家康拉德·沃丁顿(Conrad Wadding‐ton,1905—1975)为了阐明发育生物学中的相关概念提出了表观遗传景观图(图1),尝试解释胚胎发育过程中环境信号对细胞基因组的影响。表观遗传景观图形象隐喻了一个单细胞经分裂分化后形成各种类型细胞的过程。该图类似于山地间起伏的地形图,山谷不同的路径代表细胞分化的路径,谷底有不同的凹槽,受精卵类似于山顶的球,受精卵分裂后,球越来越多,便向山谷中滚动,滚动过程中遇到岔口,在不同的信号引导下,选择不同的路径,也就是分化方向不同,最终落入谷底的凹槽中,一旦进入凹槽,球便无法出来,可长久保持分化状态。但并不是所有的球最终都会滚入凹槽,部分细胞被困在半山腰,形成多能的细胞,但随时做好准备滚落山谷。表观遗传景观图强调细胞分化的命运是动态的、持续的,不是瞬间决定的,形象地解释了同一个体不同的细胞虽具有相同的DNA分子,却表现出不同的细胞命运决定的原因。康拉德·沃丁顿提出了表观遗传景观图,从现象层面描述了表观遗传。后来分子生物学的发展才逐渐揭开部分表观遗传的分子机制。
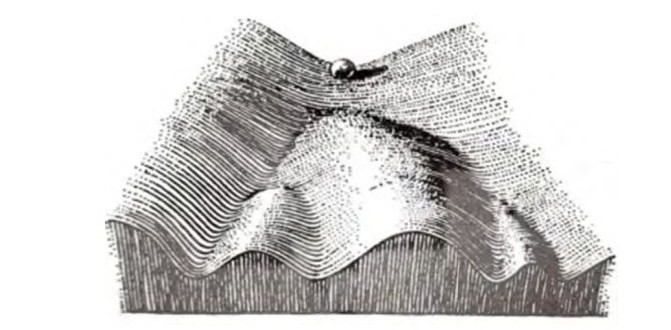
图1康拉德·沃丁顿提出的表观遗传景观图
2 细胞全能性表达的多因素调控机制
细胞分化过程受转录因子、表观遗传、环境等的共同影响。细胞分化过程中,基因序列不变,基因表达的方式改变。
细胞全能性表达的过程是基因表达调控、表观遗传修饰、微环境信号等多因素调控的结果。转录因子在基因表达调控中起关键作用。转录因子是一类能与基因启动子等调控区域特定DNA序列结合的蛋白质或RNA,可调控基因的转录。在胚胎发育过程中,不同的转录因子在特定的时间和空间顺序表达,引导细胞向不同的方向分化,形成各种组织和器官。转录因子可识别并结合基因的启动子、增强子等调控区域,调控基因的转录,转录因子能招募特定的表观遗传修饰酶,与DNA甲基转移酶、非编码RNA等相互作用影响表观遗传,而表观遗传修饰又受环境影响。表观遗传修饰主要包括DNA甲基化、组蛋白修饰、染色质重塑等方式,在不改变DNA序列的基础上,对基因表达进行调控。表观遗传标记在细胞全能性表达中动态变化,精细调控相关基因是否表达或者表达量的多少。在植物组织培养、核移植、诱导多能干细胞(iPS细胞)技术等过程中,优化培养基的成分,能够显著提高细胞全能性表达的成功率,这充分说明外部因素对于细胞全能性表达的重要性。单个植物细胞发育成植株,其自身产生的生长素对全能性的表达至关重要。内部因素中的转录因子和表观遗传修饰以及外部因素,各自通过独特的机制和相互作用对细胞全能性的表达进行整体调控。
3 植物细胞全能性表达的多因素调控
3.1表观遗传景观图视角下从现象层面解析植物组织培养过程
表观遗传景观图中,球落入不同的凹槽具有不同的命运,是由于经过了不同的山谷路径,即环境不同。虽然这个过程中基因序列并未发生改变,但是它们的表观遗传修饰不同,这些修饰虽不改变DNA序列,却能对基因的表达及表达水平产生影响。之前人们一直认为球滚落到凹槽以后便无法跳出,细胞会长久保持这种分化状态,细胞分化经过的路径是单行道,是不可逆的,因为它们受控于特定的环境。1958年,斯图尔德(Steward,1902—1972)等取胡萝卜根韧皮部的细胞离体培养后,最终培育出了一棵胡萝卜植株。这个实验证明了细胞具有全能性,也就是表观遗传景观图中滚落到凹槽的细胞可以重新回到山顶,与受精卵类似,具有发育成完整个体的能力。
基于表观遗传景观图的分析,如果把凹槽中的球取出,擦除它的表观遗传修饰,重新推回山顶,则这个球如同受精卵具有全能性,此过程就是对细胞进行了重编程。植物组织培养过程中细胞的离体,类似于从凹槽中取出球,脱分化也就是擦除表观遗传修饰后将球推回山顶,再分化则类似于球到达山顶后再次沿不同的路径分化的过程,这是细胞全能性表达的过程。这是基于表观遗传景观图,从现象层面解析植物组织培养中的有关问题。
受精卵是自然发育中具备完全全能性的细胞,依赖自然进化形成的协同调控体系,表观遗传修饰处于高度的可塑性状态。而细胞重编程是打破细胞已分化命运重新具有了全能性,依赖人工构建的补偿调控体系,细胞也需要“克服”已分化细胞的表观遗传记忆。2类细胞虽都表现出全能性特征,但是重编程细胞发育无法达到受精卵的完全的、自然的全能性,若发育中移除某些外源信号,全能性便不能表达或者发育异常。
3.2 植物细胞全能性表达的分子机制
植物体细胞在未离体的情况下,会受控于其所在的微环境信号的调控,只能形成特定的细胞。伤口是启动植物细胞重编程的重要信号,离体的组织和母体的交流无法进行,摆脱了受控的微环境,释放出很多信号启动细胞的重编程。植物在受到外伤后会进行再生和自我修复,这是植物自我保护的本能反应。
植物体细胞全能性表达,需要抑制其分化方向上细胞的特异性基因表达,激活其全能性表达的相关基因,改变细胞分化命运。2025年山东农业大学张宪省团队在对拟南芥的研究中,成功地将叶片表皮细胞重编程为全能性干细胞,再生成胚胎,最终发育成植株,细胞不经过愈伤组织阶段直接发育成个体。该团队的研究首次揭示了单个植物体细胞发育成植物体的分子机制。人工诱导高表达基因LEC2,可招募DNA去甲基化酶,特异性去除生长素合成基因启动子区域的甲基化标记,甲基化水平降低,染色质结构松散,LEC2转录因子与生长素合成基因启动子区域结合,启动相关的基因转录,细胞内生长素积累。SPCH基因是叶片气孔前体细胞中特有的基因,招募组蛋白修饰酶,维持染色体的松散状态,招募转录激活因子,生长素合成基因持续高效表达,植物细胞内自身生长素的积累为全能性表达提供稳定的分子基础,是全能性被激活的重要信号。LEC2和SPCH二者协同,激活生长素合成通路,气孔细胞脱离气孔发育之路,成为中间态,细胞中染色质重塑,全能性基因被逐步激活表达,气孔发育分化的相关基因沉默,气孔细胞获得全能性。此过程中细胞的全能性获得需要内源生长素信号通路、特定转录因子诱导和表观遗传重塑等多因素调控,确保全能性的有序表达。
4 动物细胞全能性的多因素调控
4.1 核移植中细胞全能性表达调节机制
理论上,动物细胞也具有全能性的潜能,但全能性的表达往往受到诸多限制。基于表观遗传景观图的分析,若要使动物细胞的全能性表达,需要对细胞的表观遗传修饰进行重编程。减数分裂Ⅱ期卵母细胞储备丰富的重编程因子,在精卵结合后的36 h内,卵母细胞会高效地擦除除印记区域以外的表观遗传修饰。当体细胞的细胞核移植到卵母细胞中时,卵母细胞也会对体细胞的细胞核进行表观遗传修饰的擦除,但擦除效率相对较低。这可能是因为体细胞在分化过程中,形成了更为复杂且稳定的表观遗传屏障,增加了卵母细胞擦除修饰并重置基因表达程序的难度,而精子形成过程中,经历了独特的表观遗传修饰,更容易被卵母细胞重编程机制所作用。胚胎细胞具有较高的发育潜能和分化能力,在重编程等方面的技术难度较低,因此胚胎细胞核移植成功率高于体细胞核移植。
灵长类动物的体细胞核移植存在卵母细胞去核难、胚胎发育率低等问题。2017年,我国科学家成功攻克了长期以来阻碍体细胞克隆猴诞生的难题,培育出克隆猴“中中”和“华华”。研究发现,核移植的成功率低主要原因是供体细胞核中存在重编程抗性区(RRRs),染色质和基因组上的表观遗传修饰阻碍影响了全能性表达。为移除核移植过程中胚胎发育早期的表观遗传屏障,实现对相关基因表达的精准调节,研究人员向重构胚注入组蛋白去甲基化酶Kdm4d的mRNA,同时用组蛋白乙酰酶抑制剂TSA对其进行处理。组蛋白上抑制性标记导致转录因子无法与全能性基因的启动子结合,全能性基因无法被激活。去甲基化酶Kdm4d的mRNA表达出的组蛋白去甲基化酶可擦除特定的组蛋白甲基化修饰,组蛋白去乙酰酶抑制剂TSA能提高组蛋白乙酰化水平,二者协同促进染色质结构重塑,染色质变得松散,转录因子结合到基因上,提高相关基因表达,克服了表观遗传屏障,促进囊胚的发育。
4.2 iPS细胞技术的分子机制
克隆技术已证实,体细胞核含有完整的遗传信息。2006年,日本科学家山中伸弥(Shinya Yamanaka,1962—)选择成纤维细胞作为实验材料。对筛选出的最有可能的24个基因采用不同的组合方式再次筛选,向成纤维细胞中导入Oct4、Sox2、Klf4、c-Myc这4个关键基因,成功获得iPS细胞。他的研究表明,借助这几个关键基因,就能推动细胞的重编程。动物细胞重编程,也需要抑制其分化方向上细胞的特异性基因表达,激活其全能性表达的相关基因,改变细胞分化命运。Klf4可抑制细胞的分化程序,向多能干细胞转变。Oct4基因表达的Oct4蛋白属于转录因子,能招募表观遗传酶对染色质进行修饰,调控基因表达。在体细胞中,Oct4基因通常处于不表达状态,然而,一旦体细胞中Oct4基因得以表达,这些细胞便会呈现出类似早期发育细胞的特征。Sox2可参与染色质重塑,使沉默的基因得以重新激活,Sox2与Oct4协同,共同调控激活全能性表达的相关基因,这对于重编程至关重要。c-Myc属于原癌基因,可激活大量与细胞增殖和代谢有关的基因,促进细胞周期进程及细胞向多能状态的转变。但c-Myc具有致癌性,后来常被替换或去除。后来科学家将特定的蛋白、基因、小分子化合物导入已分化的T细胞和B细胞,诱导形成iPS细胞。目前已经可以将人的成纤维细胞不经iPS细胞,直接转化为神经元细胞,这相当于把球推到半山腰就转到另一个槽里。
早期研究认为,iPS细胞不具有全能性,只有多能性。2009年,中国科学院动物研究所周琪研究组和上海交通大学医学院曾凡一研究组在世界上首次证明了iPS细胞的全能性。该发现将干细胞、发育生物学和再生医学领域的研究推到一个新的高度。
表观遗传景观图,为人们提供了开拓思维和有效思考的途径。细胞全能性的表达,涉及许多表观遗传信息的改变,因此植物组织培养和核移植技术成功的关键主要是如何正确擦除表观遗传信息。对细胞重编程,需要研究在细胞分裂、分化中什么信息引导细胞的命运决定,加入哪些表观遗传修饰。未来的研究需要继续探索转录因子、表观遗传调控网络和环境的相互作用对全能性表达的影响,提高再生的成功率,从而更好地服务生产和生活。
来源:李晓燕,孙鹏飞,王艳,等.从表观遗传景观图到多因素调控解析高中生物学细胞全能性相关问题[J].生物学通报,2026,61(3):91-94.


近期评论